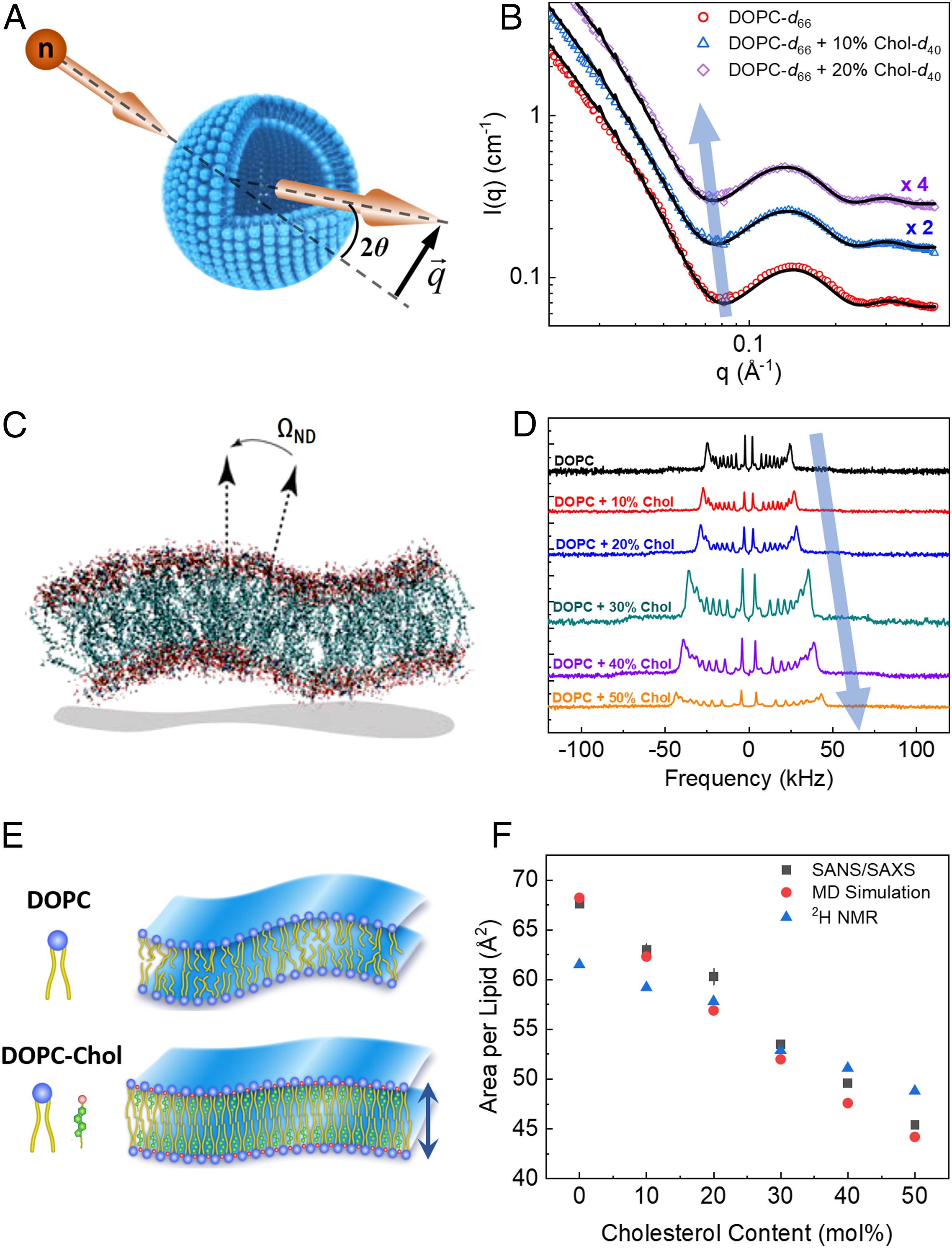
SIGNIFICANCE: Cholesterol regulates critical cell functions, including lysis, viral budding, and antibiotic resistance, by modifying the bending rigidity of cell membranes; i.e., the ability of membranes to bend or withstand mechanical stresses. A molecular-level understanding of these functions requires knowledge of how cholesterol modifies membrane mechanics over relevant length and time scales. Currently, it is widely accepted that cholesterol has no effect on the mechanical properties of unsaturated lipid membranes, implying that viruses, for example, can bud from regions enriched in (poly)unsaturated lipids. Our observations that cholesterol causes local stiffening in DOPC membranes indicate that a reassessment of existing concepts is necessary. These findings have far-reaching implications in understanding cholesterol's role in biology and its applications in bioengineering and drug design.
ABSTRACT: Cholesterol is an integral component of eukaryotic cell membranes and a key molecule in controlling membrane fluidity, organization, and other physicochemical parameters. It also plays a regulatory function in antibiotic drug resistance and the immune response of cells against viruses, by stabilizing the membrane against structural damage. While it is well understood that, structurally, cholesterol exhibits a densification effect on fluid lipid membranes, its effects on membrane bending rigidity are assumed to be nonuniversal; i.e., cholesterol stiffens saturated lipid membranes, but has no stiffening effect on membranes populated by unsaturated lipids, such as 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC). This observation presents a clear challenge to structure–property relationships and to our understanding of cholesterol-mediated biological functions. Here, using a comprehensive approach–combining neutron spin-echo (NSE) spectroscopy, solid-state deuterium NMR (2H NMR) spectroscopy, and molecular dynamics (MD) simulations–we report that cholesterol locally increases the bending rigidity of DOPC membranes, similar to saturated membranes, by increasing the bilayer's packing density. All three techniques, inherently sensitive to mesoscale bending fluctuations, show up to a threefold increase in effective bending rigidity with increasing cholesterol content approaching a mole fraction of 50%. Our observations are in good agreement with the known effects of cholesterol on the area-compressibility modulus and membrane structure, reaffirming membrane structure–property relationships. The current findings point to a scale-dependent manifestation of membrane properties, highlighting the need to reassess cholesterol's role in controlling membrane bending rigidity over mesoscopic length and time scales of important biological functions, such as viral budding and lipid–protein interactions.
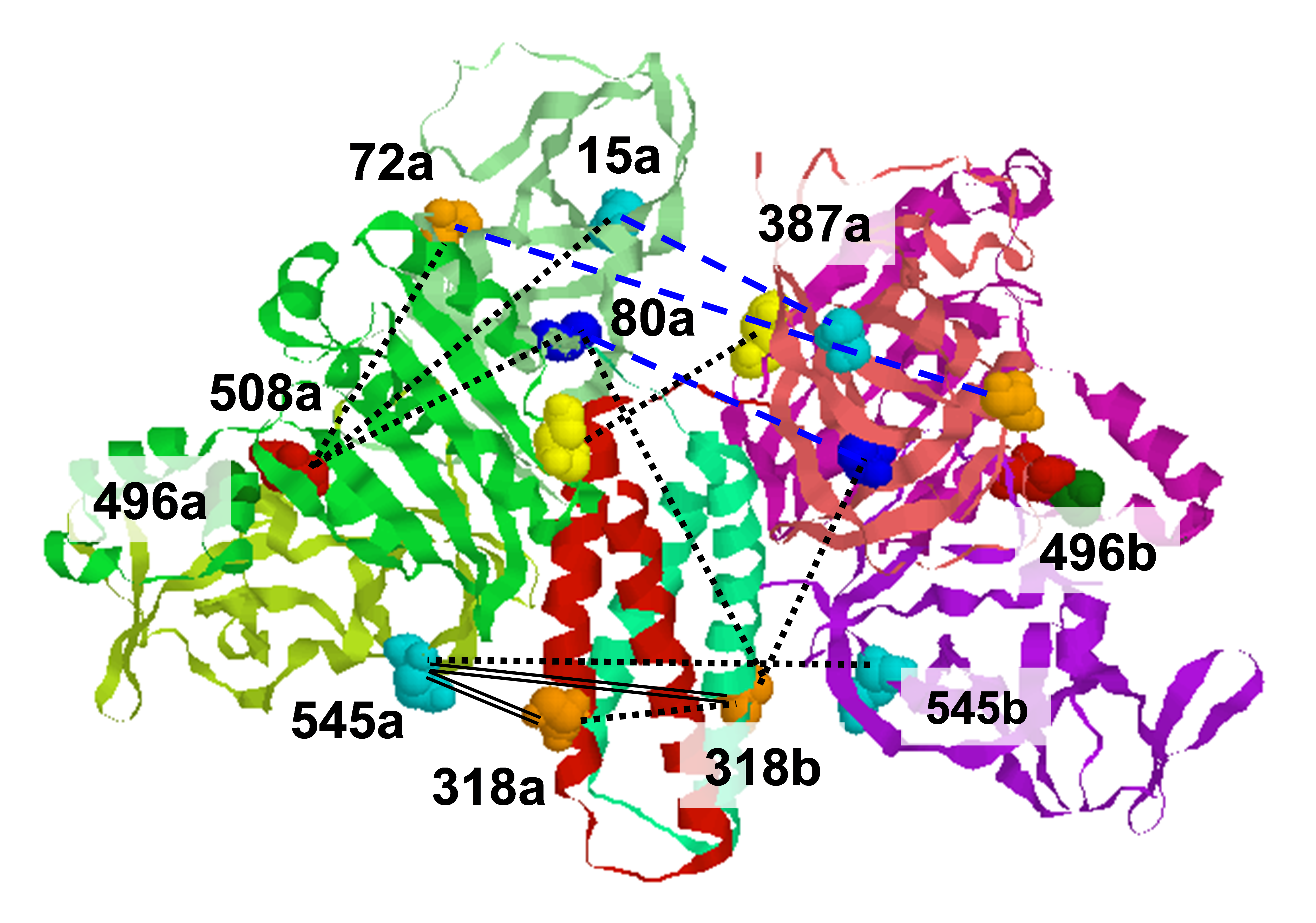
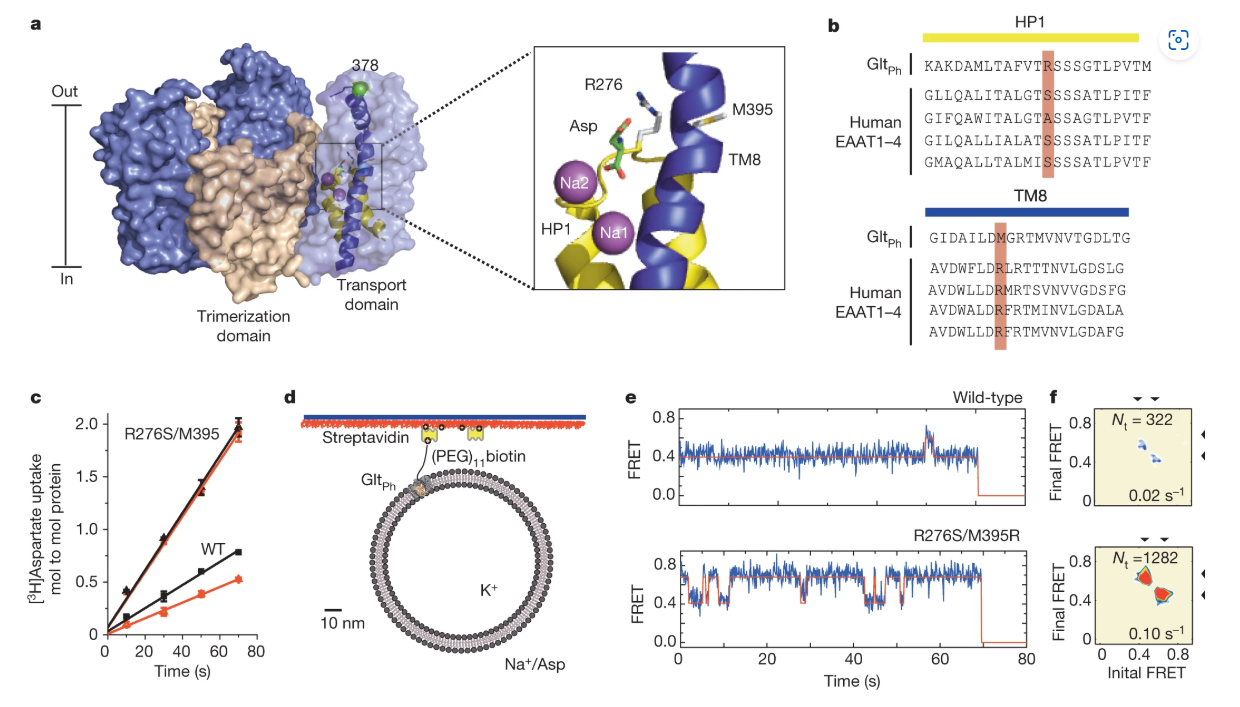
ABSTRACT: Glutamate transporters terminate neurotransmission by clearing synaptically released glutamate from the extracellular space, allowing repeated rounds of signalling and preventing glutamate-mediated excitotoxicity. Crystallographic studies of a glutamate transporter homologue from the archaeon Pyrococcus horikoshii, GltPh, showed that distinct transport domains translocate substrates into the cytoplasm by moving across the membrane within a central trimerization scaffold. Here we report direct observations of these 'elevator-like' transport domain motions in the context of reconstituted proteoliposomes and physiological ion gradients using single-molecule fluorescence resonance energy transfer (smFRET) imaging. We show that GltPh bearing two mutations introduced to impart characteristics of the human transporter exhibits markedly increased transport domain dynamics, which parallels an increased rate of substrate transport, thereby establishing a direct temporal relationship between transport domain motion and substrate uptake. Crystallographic and computational investigations corroborated these findings by revealing that the 'humanizing' mutations favour structurally 'unlocked' intermediate states in the transport cycle exhibiting increased solvent occupancy at the interface between the transport domain and the trimeric scaffold.
.svg)